
虽然杏鲍菇已经进行了大规模商业化栽培,但是关于杏鲍菇生长发育关键基因研究较少。本研究为了研究一个G蛋白α亚基基因PeGNAI在生长发育中的作用,构建了的过表达(PeGNAI-OE)和RNA干扰(PeGNAI-RNAi)突变株。研究表明GNAI的表达能够影响菌丝生长和索状联合的数量。转化子具有更高的cAMP水平,这可能参与了杏鲍菇的原基的形成。与野生型相比较7个收到GNAI调控的下游蛋白在PeGNAI-OE和PeGNAI-RNAi突变株中差异表达。这些基因可能与菌丝生长和酶活相关,涉及了MAPK信号通路、肌醇磷酸盐代谢、抗坏血酸盐、醛糖二酸盐代谢和淀粉、这趟代谢等过个通路。研究发现了GNAI基因能够通过不同的生理学过程调控杏鲍菇的生长和发育,GNAI参与的转录调控非常复杂需要进一步深入研究。
cAMP途径在植物、真菌和细菌细胞内信号转导中起到了非常重要的作用。Heterotrimeric guanine nucleotide-binding proteins (G proteins)在cAMP途径中扮演了重要角色。G proteins由ɑβγ三个亚基构成,根据序列和功能功能G蛋白ɑ亚基可以分成四个家族 Gαs, Gαi, GαQ, andGα12。真菌中G蛋白能够被G蛋白偶联受体(GPCRs)所激活,随后G蛋白激活下游的信号转导过程包括adenylate cyclase (AC), mitogen-activated protein kinase (MAPK)s, ion channels, phosphodiesterase, and phospholipase。真菌中G蛋白控制了很多生理过程例如营养生长、分生孢子形成、抗逆、环境和营养响应等。有研究表明Gα亚基能够影响Metarhizium robertsii分子生包子萌发,细胞壁形成和抗热有关。在Phytophthora sojae中Gα亚基可以与下游的PsYPK1相互作用,的缺失会造成孢子囊和卵孢子的减少从而降低菌丝生长速度。在Ciboria shiraiana中沉默两个Gα亚基,CsGPA1和CsGPA2,对菌丝生长没有影响但是能够降低菌核的数量并增加单个菌核的重量。在Aspergillus fumigatus中敲除gpaB和gpnA能够降低菌丝体生长速率,但是在gpaA基因敲除后菌丝生长量增加。但是杏鲍菇中G蛋白的功能仍不清楚。在之前的研究中,通过转录组研究表明cAMP信号通路和GNAI基因可能与杏鲍菇生长发育有关。
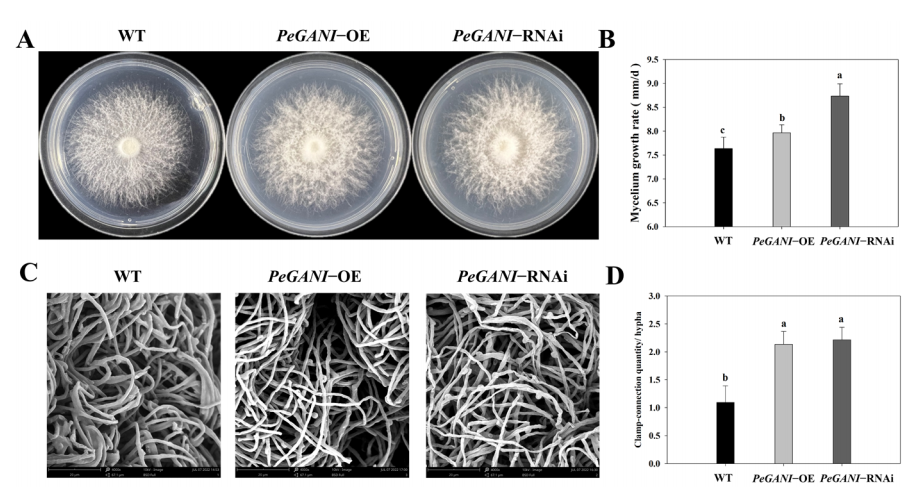
图1 野生型和转化子的菌丝生长情况
本研究中利用载体pCAMBIA1301-Hygro和农杆菌(EHA105)侵染法对GNAI基因(KAF9491299.1)进行了过表达和RNA干扰。RT-qPCR实验采用β-actin基因作为参考基因。qPCR结果显示GNAI基因在PeGNAI-OE转化子中上调了4.37倍,在PeGNAI-RNAi转化子中下调了0.65倍。将9mm的接种块接种到平板中央,通过测量直径来判断菌丝生长速率。图1展示了突变株的菌丝平板照片,可以看出来转化子倾向于产生絮状的菌丝体,中间部分菌丝体较厚,边缘部分薄。与野生型相比较,PeGNAI-RNAi转化子气生菌丝更壮,菌丝更浓密。与野生型相比较过表达(PeGNAI-OE)和RNA干扰(PeGNAI-RNAi)突变株生长速率提高了4.46%和14.42%。在接种培养7d之后,在边缘切下菌丝,浸泡到2.5%戊二醛溶液中进行固定,并进行扫描电镜观察。扫描电镜也确认了这一点。锁状联合的数量PeGNAI-RNAi>PeGNAI-OE>WT。
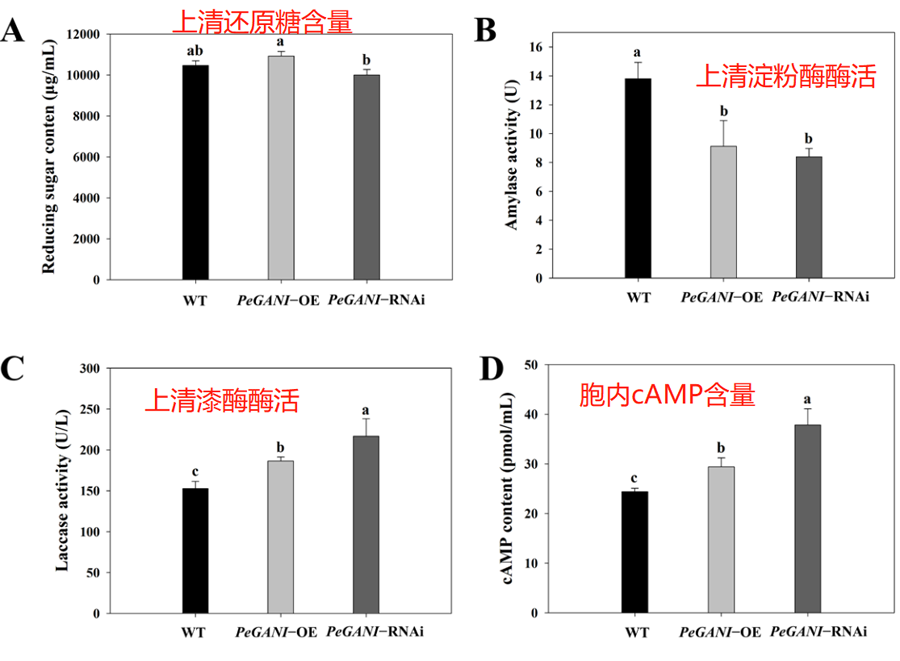
将平板上培养的菌丝块(1cm直径)接种到100 mL液体PDA培养基中,25℃培养10d,离心过滤掉菌丝,上清测定酶活。DNS法测定上清中的还原糖。ABTS法测定漆酶酶活。淀粉酶酶活根据DNS法测定产生的还原糖数量来确定。胞内的cAMP的浓度使用金瑞斯生物科技的cAMP ELISA Detection Kit进行测定。具体的结果如图2所示。
葡萄糖含量、淀粉酶、漆酶活力与菌株降解大分子化合物和营养物质利用有关系,这些都跟菌丝生长密切相关。Hu等人在研究Chaetomium globosum的时候发现,将编码Gα的基因gna1敲除之后能够阻断Gα-cAMP/PKA信号通路。C. shiraiana的研究中发现将CsGPA2沉默之后过氧化物酶和漆酶活力上升了一点,但是CsGPA1沉默之后菌株毒性下降,漆酶和过氧化物酶活力没有影响。本研究中在RNAi和过表达突变株中漆酶活力均有提高。这可能促进了菌丝对于底物的利用,从而增加了菌丝生长速率。cAMP水平在细胞相应外部刺激的时候起到了关键作用。Miwa等人的研究表明将一个G蛋白偶联受体敲除之后,突变株能够通过cAMP信号通路来影响菌丝的形态和菌丝的形成。跟我们的结果一致,Yao等人也发现在平菇PC9中转入Gα基因之后能够提高漆酶活力以及细胞内的cAMP水平。RNAi突变株中可能是因为GNAI的下调导致了腺苷酸环化酶活性下降从而使得cAMP水平升高。在过表达突变株中cAMP水平的提高可能与AC的hyperactivation or hypersensitization有关系。此外文章认为GNAI可能会通过cAMP水平调节来影响漆酶和淀粉酶的活性。
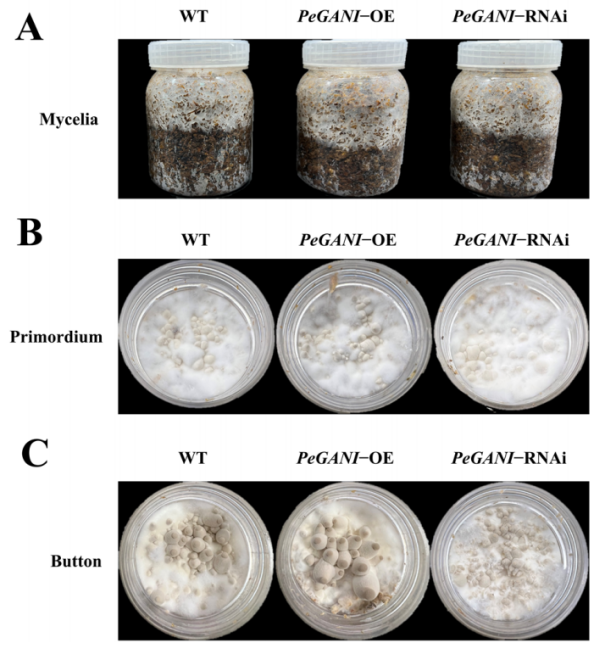
图3 组培瓶中三个菌株出菇情况
组培瓶中固体栽培料生长实验表明,过表达突变株生长速率显著高于野生型,RNA干扰株无明显差别。过表达突变株纽扣期(钉头期)出现时间更早,纽扣子实体出现的少,涨势好。RNA干扰突变株纽扣期出现最晚,原基多涨势差。可以看出来PeGNAI在杏鲍菇生长发育中起到了促进的作用。
食用菌发育主要分为营养生长和生殖生长两个过程,在两个过程中间是原基期,也是营养生长转为生殖生长的关键时期。裂褶菌的研究表明了G蛋白偶联受体与裂褶菌生长发育有关系,并且发现了一个G蛋白信号分子Thn1可能参与了裂褶菌生长发育。本研究结果也证明了G蛋白参与了杏鲍菇的生长发育过程,包括影响了菌丝积累、原机形成和钉头状子实体的数量。
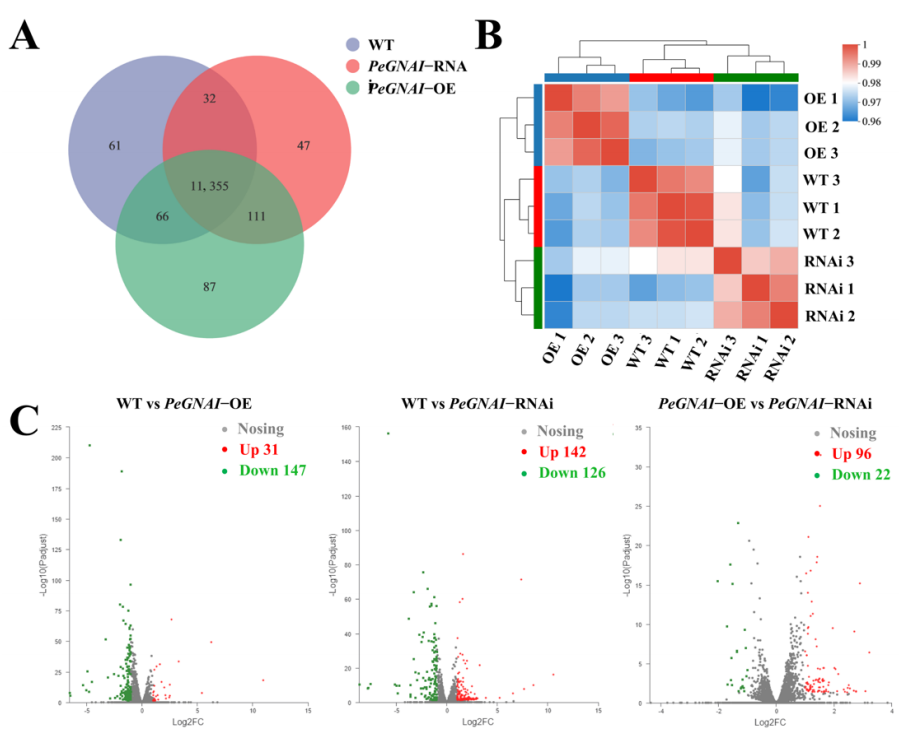
图4 转录组差异表达分析
利用Illumina Novaseq 6000平台进行了转录组测序,利用FPKM方法进行基因差异表达分析,FDR ≤ 0.05、log2FC ≥ 1作为差异表达基因判断标准。样本之间的差异采用了One-way ANOVA的方法来进行分析。相关性分析表明三个样本的相关性非常高,0.96以上。与过表达突变株相比较,WT菌株中31个基因上调,147个基因下调。与RNAi突变株比较,WT菌株中142个上调,126个下调。这表明了GNAI可能是一个负调控基因表达的转录因子。
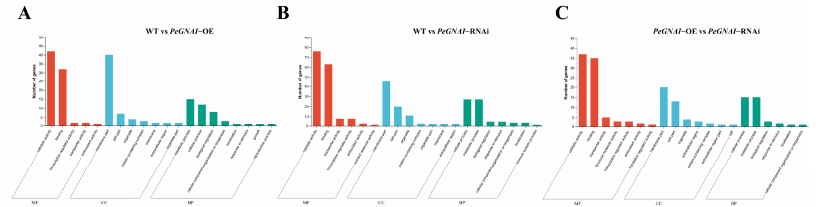
对差异蛋白的GO聚类可以看出来BP主要集中在代谢过程。因此GNAI主要通过细胞代谢来调控菌丝生长过程。CC term主要聚类到了细胞膜、细胞和细胞器分类上,说明了GNAI能够影响细胞膜的结构从而影响菌丝生长。MF term主要聚类到了binding、catalytic activity、transcription regulator activity和transporter activity,这表明GNAI可能通过调控不同酶的酶活来影响生长。

KEGG聚类分析表明,与过表达突变株相比较,野生型的差异表达基因主要聚类在MAPK信号通路、amino sugar nucleotide sugar metabolism和内吞作用上面。野生型与RNAi突变株的差异表达基因主要聚类在inositol phosphate metabolism、ascobate and aldarate metbaolism、MAPK信号通路上面。过表达和RNAi突变株的差异表达基因主要几种在methane metabolism、ascorbic acid、aldarate metabolism、pyruvate metabolism、Arginine and proline metabolism、辅因子合成上面。
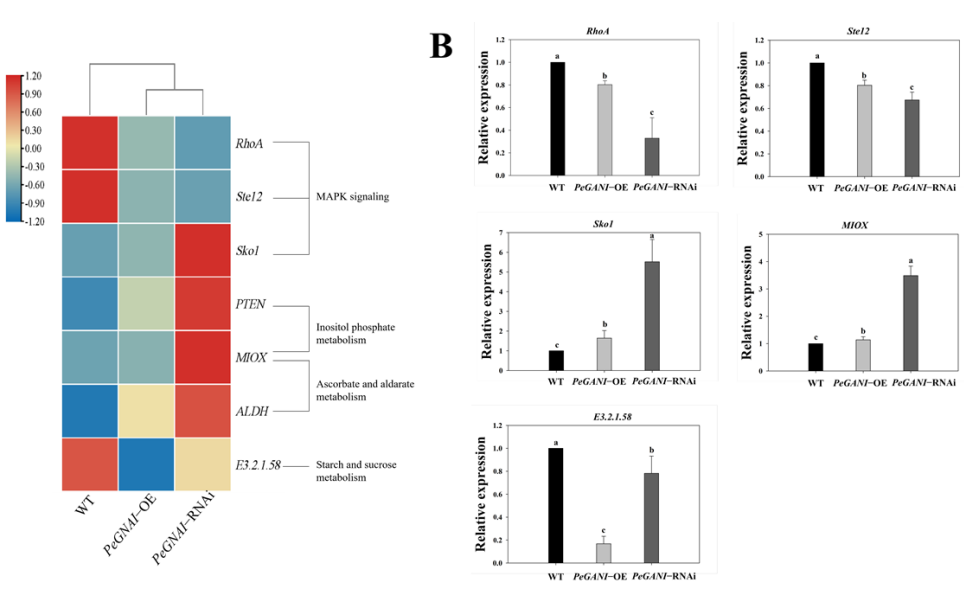
进一步本研究对于菌丝生长发育的一些保守的通路的关键基因进行了深入分析。与WT相比较MAPK信号通路的相关基因(RhoA和Ste12)在过表达突变株中下调。与WT相比较,MAPK信号通路相关的Sko1基因表达量上调。与inositol phosphate metabolism相关的表达基因MIOX和PTEN在RNAi突变株中均显著上调。跟ascobate and aldarate metbaolism相关的MIOX和ALDH基因在RNAi干扰突变株中也上调了。q-PCR的结果与转录组结果一致。
在真菌生长发育过程中MAPK途径扮演了很重要的角色,其中在真核生物中small monomeric GTPases作为分子开关调控了很多细胞过程。Small GTPases主要分成了5个家族,其中Rho家族是研究最为深入的一个家族,其中真核生物中RhoA、Rac1、Cdc42研究的比较多。Ras GTPases在丝状真菌的孢子萌发、菌丝分支、无性繁殖中扮演了重要的角色。Talaromyces pinophilus菌株中敲除了seb1基因后,造成了形态缺陷和分支增多。本研究中在过表达和缺失突变株中均发现了RhoA基因的下调,这可能是造成了突变株菌丝更加茂盛的原因。酿酒酵母中研究中发现转录因子Ste12是MAPK Fus3/Kss1的直接下游靶基因。Dolan等人发现Ste12能够抑制丝状真菌菌丝的生长。并且研究还发现Ste12可能跟致病性有关系。本研究中发现突变株中Ste12下调,可能跟菌丝的快速生长有关系。研究表明加速酵母中Sko1与菌丝生长和压力胁迫响应密切相关,将Sko1敲除后能够使得假丝酵母固体培养上菌落形态改变,液体培养基中更容易形成菌丝。杏鲍菇中的RNAi突变株中Sko1基因表达量上调可能也会通过MAPK途径影响菌丝生长。
PTEN是一个Tensin-like phosphatase 1 (TEP1),在禾谷镰孢菌中敲除FgTEP1会造成菌丝体对lithium敏感,并减少分生孢子的数量。而TEP1缺失的酵母突变株对lithium and wortmannin具有抗性。MIOX基因与糖分解过程有关系,具有inositol oxygenase活性,ALDH具有ATP结合和蛋白激酶活性。在RNAi突变株中PTEN、ALDH和MIOC基因表达量都显著上调,这说明了GNAI沉默能够增加蛋白磷酸酶和inositol氧化酶活力,促进ATP与某些蛋白结合,从而激活PKC,后者会通过cAMP信号通路调节杏鲍菇的生长发育。
与starch and sucrose metabolism相关的E.3.2.1.58基因在突变株中表达量均下调。前面的实验也证明了突变株的淀粉酶活性下降,这与转录组和qPCR结果一致。Hu等人再研究Chaetomium globosum的时候发现,敲除了pG14(Gα基因gna1)之后与淀粉和蔗糖代谢相关的通路中46个下调,只有2个上调。这跟我们的实验结果是一致的。
G-protein α subunit如何影响下游基因的表达需要进一步深入研究,比如通过蛋白相互作用研究来验证。本研究为G-protein α subunit在食用菌菌丝生长、有性繁殖和子实体发育中的功能研究提供了重要参考。
qPCR Ref基因序列:
β-actin-F:GCTGGTATCCACGAGACAACA
β-actin-R:CAAGATAGAACCACCAATCCAAA
>KAF9491299.1 guanine nucleotide binding protein, alpha subunit [Pleurotus eryngii]
MGCVQSSGVDDEAKARNDEIENQLKRDRMMAKNEIKMLLLGAGESGKSTVLKQMKLIHHG
GYNEQERDSYKEIIFSNTIQSMRAILEAMPQLDISLAPQNDARRAIIMALPLQVEVDILP
RDVGDAIRGLWRDPGVKEAVRRSREFQLNDSAIYYFNAIDRMSSPGYMPTDQDILRSRVK
TTGITETTFKVGELTYKLFDVGGQRSERKKWIHCFENVTALVFLVSLSEYDQMLYEDESV
NRMQEALTLFDSICNSRWFVKTSIILFLNKIDLFAEKLPRSPLGDYFPDYTGGDSYDAAC
NYLLNRFVSLNQSAATKQIYAHYTCATDTQQIKFVLSAIQDILLQLHLRECGLL
附件1: 点击查看pdf