
茶树菇能够利用多种底物进行生长,本身具有多种CAZymes,营养类型介于典型的白腐菌和腐殖质分解者之间。由于木头里面的N元素较少,因此茶树菇会从周围的植物凋落物(plant litter)里面吸收N元素。外加N源对于实验室条件下茶树菇生长发育的影响尚无研究。本研究中,我们通过外加β-adenosine(一种植物凋落物中的常见N源)。通过增加外源β-腺苷的水平能够显著增加气生菌丝的干重,增加成熟子实体的干重,增加原基产生的数量,并且能够显著缩短原基形成的时间。研究结果表明茶树菇能够将外加的N元直接转化为营养生长和生殖生长的生物质。在自然环境中茶树菇更倾向于在富含N源的环境中生存,因此由于人类活动导致的世界范围内N源的改变可能会影响菌丝和子实体的形成以及响应的食物网。
为了更好的控制实验参数,防止复杂培养基例如秸秆、木屑、谷物作为底物的混合培养基,对结果造成的影响,本研究的实验都在琼脂糖培养基中完成。出菇是在1.5%的MEA平板上完成的。添加两种浓度的β-腺苷,1x浓度为15 μg/mL,10x的浓度为150μg/mL。MEA经过高压蒸汽灭菌后温度降到40℃的时候,将过滤除菌的β-腺苷加入到平板上面。
从1.5%MEA平板上培养的菌丝边缘取0.3cm的菌丝体加入到不同的MEA平板上面,保留一个正常培养,其余的用铝箔包裹,所有的平板都在25℃培养,等没有包裹铝箔的对照平板长满平板(双核菌丝~13 day)之后,将平板转移到出菇条件。将平板放到70%乙醇灭菌的出菇盒中,在盒子底部铺上2个加入了10 mL无菌水的吸水纸,每个盒子里面放3个平板。为了进一步刺激菌丝在平板上面出菇,从每个平板上面去掉一个直径为0.3 vm的菌块(机械损伤),平板上盖去掉后放到盒子底部。出菇盒放到20摄氏度人工气候箱中,12h光照、12h黑暗。光照强度为10 ± 0.5 µmol m−2 s−1。接种后第21-23天就可以看到双核菌丝的子实体了,单核菌丝会更晚一些。
1.5 MEA:malt extract agar,1.5% malt extract(70167-500G,Sigma-Aldrich Chemie GmbH Munich),1.5% agar。
β-adenosine:Carl roth GmbH & Co. KG, Karlsruhe, No. A9251-5G。
Domondon在2004年有一篇文章【10233,10.1016/j.phytochem.2003.11.004】报道,β-adenosine是grass chaff中的一种活性成分,能够刺激蘑菇的产生。通过向秸秆中添加30%的Lolium perenne grass chaff能够显著提高秀珍菇和大球盖菇的产量,进一步通过纯化发现了β-adenosine是主要的活性成分。通过向1kg秸秆中添加12 mg β-adenosine就可以显著缩短秀珍菇、大球盖菇原基产生的时间。当加入25 mg/kg秸秆的β-adenosine的时候,秀珍菇产量提高了52%,大球盖菇产量提高了258%。

图1 β-腺苷结构、出菇盒和平板出菇照片
实验通过测量菌丝体的直径来判断菌丝体的生长速度,结果表明添加β-腺苷之后菌丝体生长速率并没有显著差异(图2a)。
在菌丝长满平板的时候,小心的将菌丝体刮下来,放到离心管中40摄氏度烘干24 h,称干重。结果显示加入了β-腺苷之后菌丝干重显著增加(图2B)。
实验测量了从刺激出菇开始到第一个原基产生的时间,从图2C中可以看出来,添加了β-腺苷之后从诱导出菇开始到第一个原基产生的时间显著缩短了。不加β-腺苷平均需要9.7d才能产生原基,加入1x的β-腺苷平均时间为9.2 day,加入了10x的β-腺苷之后时间缩短到了8.1 day。
此外我们还测量了下面这些参数,原基产生的数量,从刺激出菇开始到原基产生的时间,成熟子实体的干重。
从诱导出菇开始40天内所有的新产生的原基的数量都进行了统计。从结果也可以看出来,加入了β-腺苷之后产生的原基的数量显著增加(图2D)。
实验还测量了成熟子实体的干重,成熟子实体的判断标准是开始释放孢子,所有的成熟子实体从菌柄的最底部切除,转移到空的平板上40℃烘干24 h,测量干重。结果显示加入了β-腺苷之后成熟子实体的干重显著增加(图2E)。
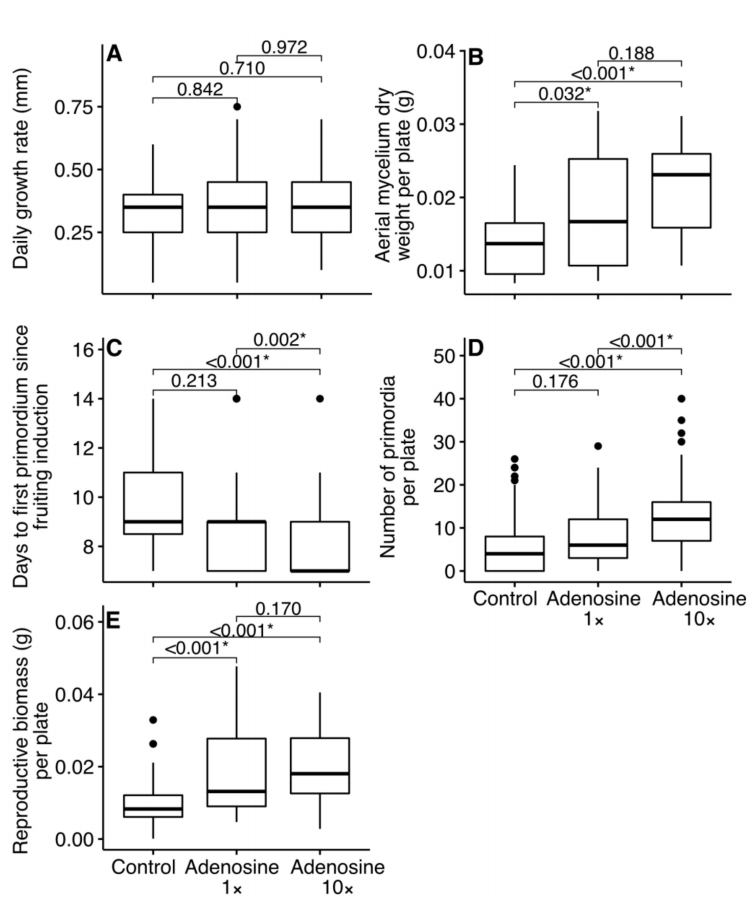
图2 添加β-腺苷对于茶树菇平板出菇的影响。
Domondon等发现添加了30%的ryegrass extract能够加快大球盖菇菌丝生长速度,但是秀珍菇的生长速度不受影响。我们的研究中显示添加β-腺苷并不会对茶树菇的菌丝生长速度产生影响。Domondon等人观察到了外加β-腺苷能够增加秀珍菇菌丝体的密度,如果计算干重的话干重也会提高。Zhang等人观察到当添加了32 μg/mL的β-腺苷之后与16 μg/mL腺苷相比较,干重出现了下降,但是在本次的研究中并没有发现生物量下降的现象。
Domondon和本研究都观察到了添加β-腺苷能够缩短原基产生的时间,尤其是添加10x的β-腺苷的时候,本研究发现平板上茶树菇原基产生的时间从~10天,缩短到了~8天。Suillus luteus并没有产生原基,这可能主要是因为缺少共生的植物根导致缺少其他的刺激条件。
本研究发现添加β-腺苷能够提高子实体的生物量。Domondon等人报道只有在添加了25μg/mL的β-腺苷才能增加秀珍菇的生物量,添加12μg/mL的β-腺苷则没有效果。而且Domondon等人的结果显示添加了25μg/mL的β-腺苷会推迟秀珍菇原基产生的时间。而在本研究中10x的β-腺苷也能够加速原基产生的速率。
作者认为本研究还有很多不足,比如说应该尝试更多的有机和无机N源,观察茶树菇对于不同N源的选择。比如说测试尿素和铵盐对于菌丝生长的影响,这些在其他的研究中已经有报道。我们之所以选择β-腺苷是因为该化合物广泛存在与自然界的植物凋落物中,因此结果比较接近于自然界中茶树菇的营养获取方式。我们本研究中采用的是MEA培养基,后续实验可以尝试其他的培养基,例如ScMM培养基。
ScMM:(1L) 20 g glucose, 1.5g L-asparagine, 0.12 mg thiamine-HCL, 1 g K2HP4, 0.46 g KH2PO4, 0.5 g MgSO4·7H2O, 5 mg FeCl3·6H2O, trace elements(参考下表).

本研究从实验室水平研究了外加N源对于茶树菇生长发育的影响,结果显示茶树菇能够利用多余的N源促进自身的生长和发育过程。这对我们研究自然界中真菌如何利用营养物质并用于自身的繁殖具有重要的意义。
附件1: 点击查看pdf