
本文系Food Science and Human Wellness原创编译,欢迎分享,转载请授权。

Introduction
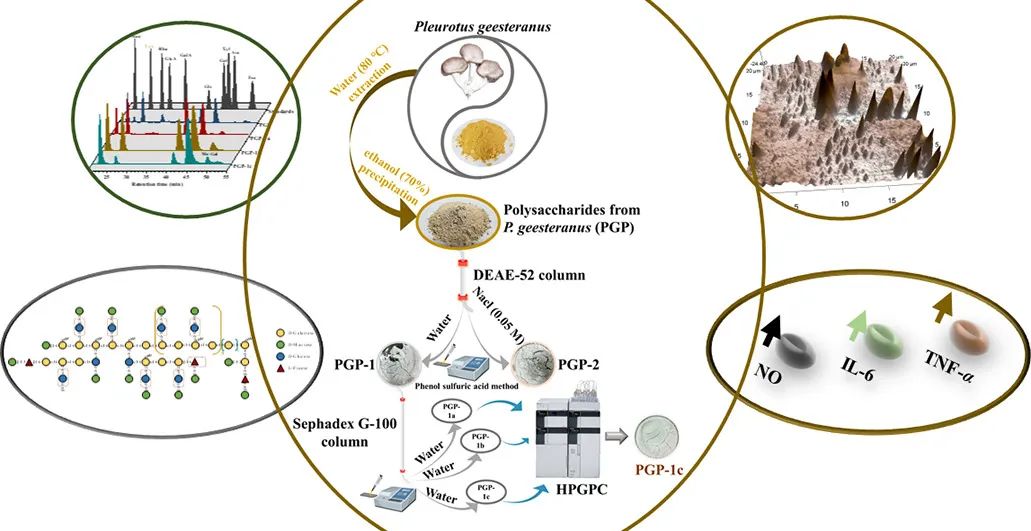
近年来,食用菌多糖的精细结构表征已成为将关键研究成果转化为免疫调节剂等功能产品的重要前提,因为多糖的生理活性与其化学成分和结构构象密切相关,这一点近年来越来越受到关注。然而,由于其结构的复杂性和多样性,对食用菌多糖结构的认识仍有很大的提升空间。侧耳科的年产量约为900万吨,是世界上最经常养殖和消费的食用菌之一。秀珍菇因其丰富的营养、优质的蛋白质和宜人的风味,已成为一种具有巨大市场价值和发展前景的食用菌。它也被认为是一种有价值的药用真菌,其多糖具有抗肿瘤、降血脂、抗氧化和保护肝脏的巨大潜力。尽管秀珍菇多糖具有巨大的药用和工业潜力,但现有的初级结构数据不足以阐明其分子机制和结构—功能关系。富含免疫调节功能多糖的食用真菌是被高度评价为免疫反应调节剂的产品之一。因此,使用食用真菌多糖为预防病毒感染的严重副作用提供了一种创新策略。虽然目前有文献证明一些来自侧耳属的多糖具有免疫活性,但目前还没有关于秀珍菇多糖免疫活性的研究。
因此,本研究对PGP-1c的结构、链式构象和免疫活性预测进行了详细描述,作为阐明秀珍菇多糖的结构—功能关系和功能产品的技术开发的第一步。
Results and Discussion
PGP-1c的糖苷联结分析
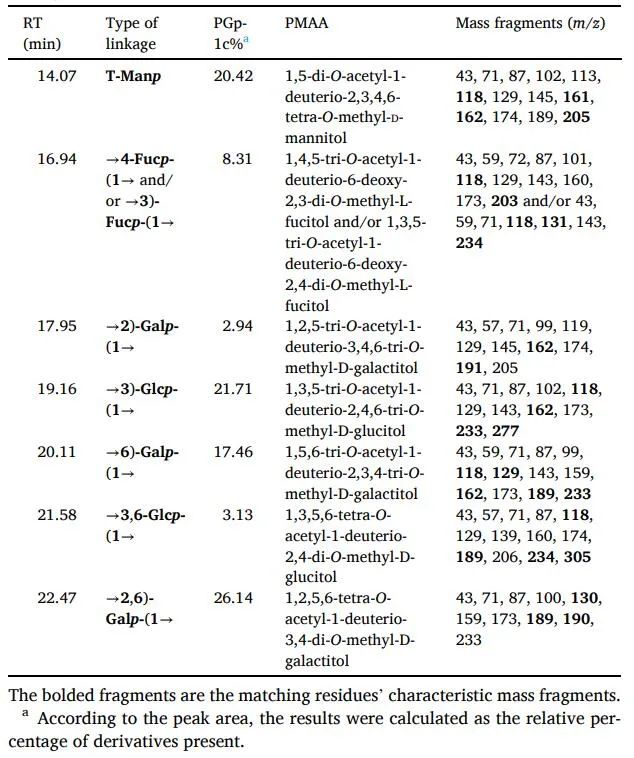
PGP-1c的甲基化分析结果见表1。在PGP-1c中发现了七个主要的连接点。主要的残留物种类是半乳糖(46.5%),其中包括→2,6)-Galp-(1→(26.1%),→6)-Galp-(1→(17.4%)和→2)-Galp-(1→(2.9%)。其次是葡萄糖(24.8%)、T-Manp(20.4%)和少量的Fuc(8.3%)。由于总离子色谱中岩藻糖的分辨率不理想,所以GC-MS无法确定→3-Fucp-(1→的量。然而,根据PGP-1c的核磁共振谱和相关文献,可以区分出→4-Fucp-(1→和→3-Fucp-(1→。该结果与单糖组成结果相互验证。此外,PGP-1c的分支度(DB)为48.6%,计算公式为DB=(NT+NB)/(NT+NB+NL),其中NT、NB和NL分别反映末端、分支和线性残基的比例含量。DB的结果证明PGP-1c具有一定的分支,核磁共振谱进一步证实了该结构。
表1 基于甲基化分析的PGP-1c联结模式
PGP-1c的核磁共振光谱分析
在1H NMR谱中确定了异构质子的化学位移范围(δ 4.50~5.39)(图1A),在δ 4.50、4.52、4.55和4.82处有异构质子信号的片段的3J1,2均大于>4,确认为β构型,质子信号>5.0的7个信号的3J1,2均<4,确认为α构型,与FT-IR的结论一致。此外,在δ 1.24和1.33的两个甲基信号被确认为单糖组成后的岩藻糖的H-6。根据1H NMR谱上四个重叠峰的积分面积,进一步结合13C NMR(图1B)、HSQC(图1D和F)和甲基化分析结果,推断出在δ 3.47/59.1和3.45/58.9的信号为O-甲基。对应于δ 5.13/101.49和5.01/100.2的异构体信号被分配为→2,6)-α-D-(3-O-Me)-Galp-(1→(残留物d)和→6)-α-D-(3-O-Me)-Galp-(1→(残留物f)。此外,β-D-Manp-(1→(残基g)的异构体信号被作为溶剂的D2O的信号峰隐藏在δ 4.82/104.5处。同样,HSQC光谱中的其他异构体信号(包括已识别的)包括5.39/102.21、5.18/103.29、5.15/101.39、5.03/100.95、5.03/100.95、4.52/105.59、4.55/105.75、4.50/105.83被推测为→3)-α-D-Glcp-(1→(残基a)、→2,6)-α-D-Galp-(1→(残基b)、→2,6)-α-D-Galp-(1→(残基c)、→6)-α-D-Galp-(→(残基e)、→6)-α-D-Galp-(1→(残留物e'),→3,6-β-D-Glcp-(1→(残留物h),→4)-β-L-Fucp-(1→(残留物i),→3)-β-L-Fucp-(1→(残留物j)。
此外,根据COSY(图1C),残留物a的H-2、H-3、H-4、H-5和H-6的化学位移分别为δ 3.71、3.95、3.69、4.03和3.90/3.76。2C)光谱,相应的13C核磁共振信号在HSQC(图1D)光谱中被分配为δ 73.4(C-2)、79.9(C-3)、75.7(C-4)、74.5(C-5)和63.81(C-6)。因此,残基a被认为是→3)-α-D-Glcp-(1→,这与单糖组成和甲基化相互印证,并与文献中的化学位移一致。其他糖残基的H-2/C-2-H-6/C-6的化学位移也是以类似的方式得出和分配的,详细的化学位移在表2中列出。值得一提的是,由于信号的重叠,PGP-1c中的一些单元和序列无法确定。结合HSQC-COSY,发现除11个残基外,在β-D-Manp-(1→被D2O隐藏的一些信号没有列在表中。但根据甲基化结果、残基的相对比例和文献记载,基本可以确认这部分未分配的信号也属于β-D-Manp-(1→。
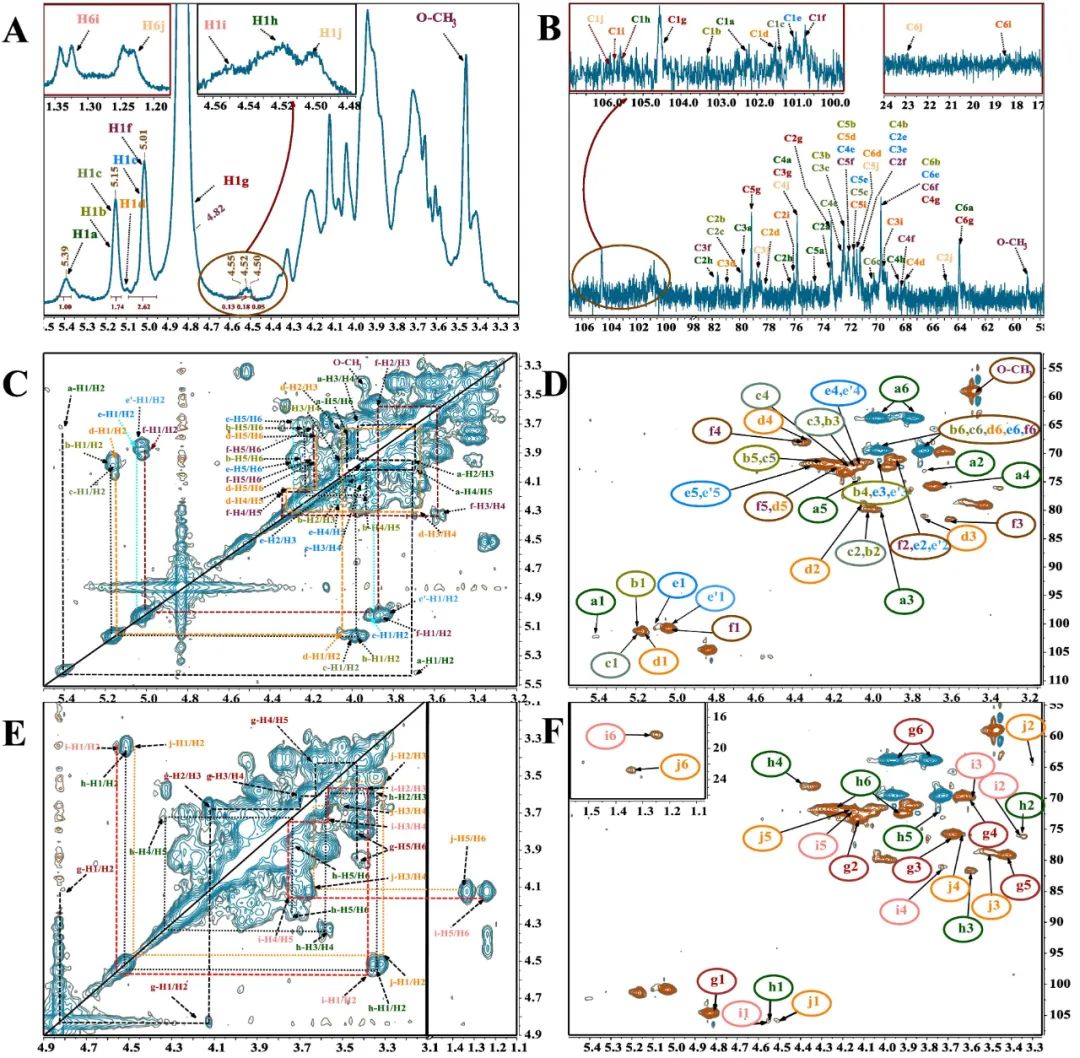
图1 PGP-1c的一维和二维核磁共振波谱
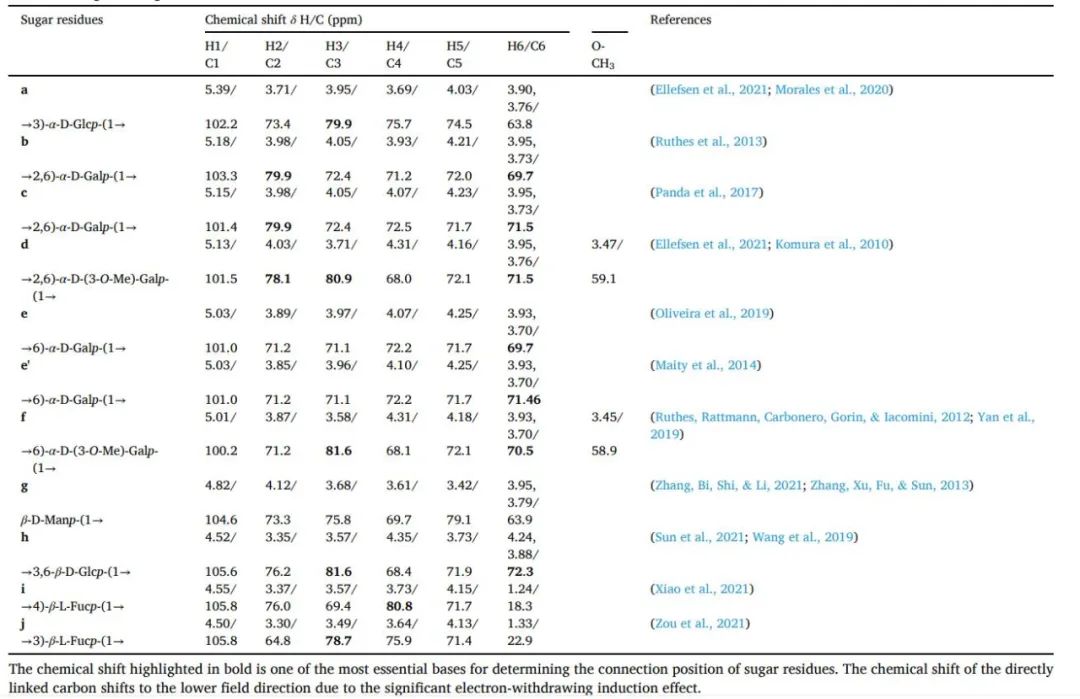
表2 PGP-1c的13C和1H信号分配
基于上述,图2描述了PGP-1c的预测模型结构,图中所示的链条构象进一步支持了这一预测。
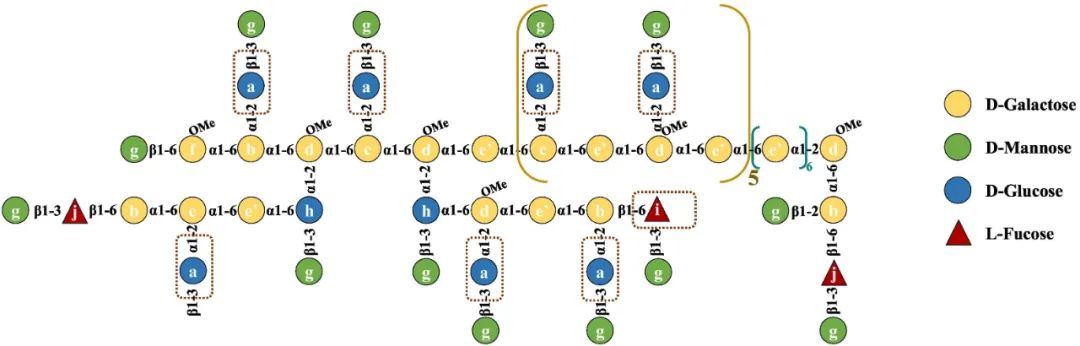
图2 PGP-1c的预测模型结构
PGP-1c的链式构象
原子力显微镜(AFM)是一种强大的技术,可以在模拟自然环境的条件下直接观察单个大分子在亚纳米尺度的构象,检测复杂体系中分子间的相互作用。较高的浓度会产生分子间的聚合甚至交联,这与分子的相互作用有关。为了研究分子形态,选择了质量浓度较低(10 μg/mL)的PGP-1c。在图3中,PGP-1c的三维结构为不同大小(高度10~100 nm,平均高度24.5 nm,宽度100~800 nm,长度300~1500 nm)的起伏。峰高可以看作是分子链高度的反映,表明支链的长度,而峰宽反映了这些分子的分子间聚集行为(不排除针尖引起的“增宽现象”)。在多糖的链状构象中,作为岛状结构的多糖并不少见,而单链的高度一般为0.1~1 nm。在杏鲍菇和平菇多糖中存在着分支和缠结的分子链,表明其形成过程不是干燥和脱水过程中出现的简单缠结或叠加,而是由于分子间的相互作用(范德华力和氢键)。可以推测PGP-1c具有长支链或叠加的多支链和一定的分子聚集,这将支持上述结构模型(图2),并表明它可能具有三螺旋结构。
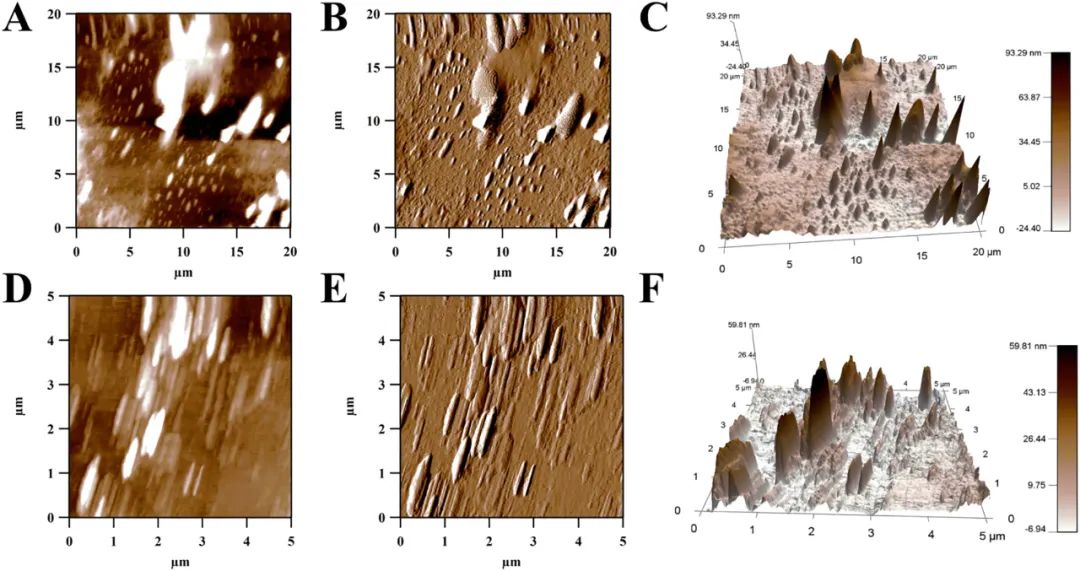
图3 PGP-1c的AFM高度(A,D),振幅(B,E)和3D(C,F)图像
XRD进一步验证了其结构特征(图4A)。在PGP-1c中,在14.7°、17.4°、30.1°、31.0°和42.0°(2θ)有不同的小肩峰,但整体上有圆形的宽峰,这表明PGP-1c中同时存在结晶和非结晶结构。具有良好对称性、无分支或少分支、强分子间力的大分子容易结晶。无定形区域的分子间力相对较弱,结晶度的增加也将增加黏度和链式聚合。
使用刚果红法发现了PGP-1c的三螺旋结构。在图4B中,PGP-1c在刚果红溶液中的存在引起了可见的最大吸收波长的向红移,表明PGP-1c-刚果红复合物已经形成,随后的结果显示,PGP-1c-刚果红复合物在低NaOH浓度下具有三螺旋结构。随着NaOH浓度的增加,PGP-1c-刚果红复合物的下降幅度大于刚果红,三螺旋构象转变为单螺旋构象,证实了AFM的结论。来自侧耳真菌的多糖经常表现出这种构象。
图4D显示了1000和5000倍放大的PGP-1c的表观结构,它富含纤维丝,有少量无序交织的带状物和一些圆形节点,形成了松散排列的网络结构。形态学研究结果与AFM相吻合,进一步证实了PGP-1c中的缠结和分支结构。这种结构与杏鲍菇多糖相似。
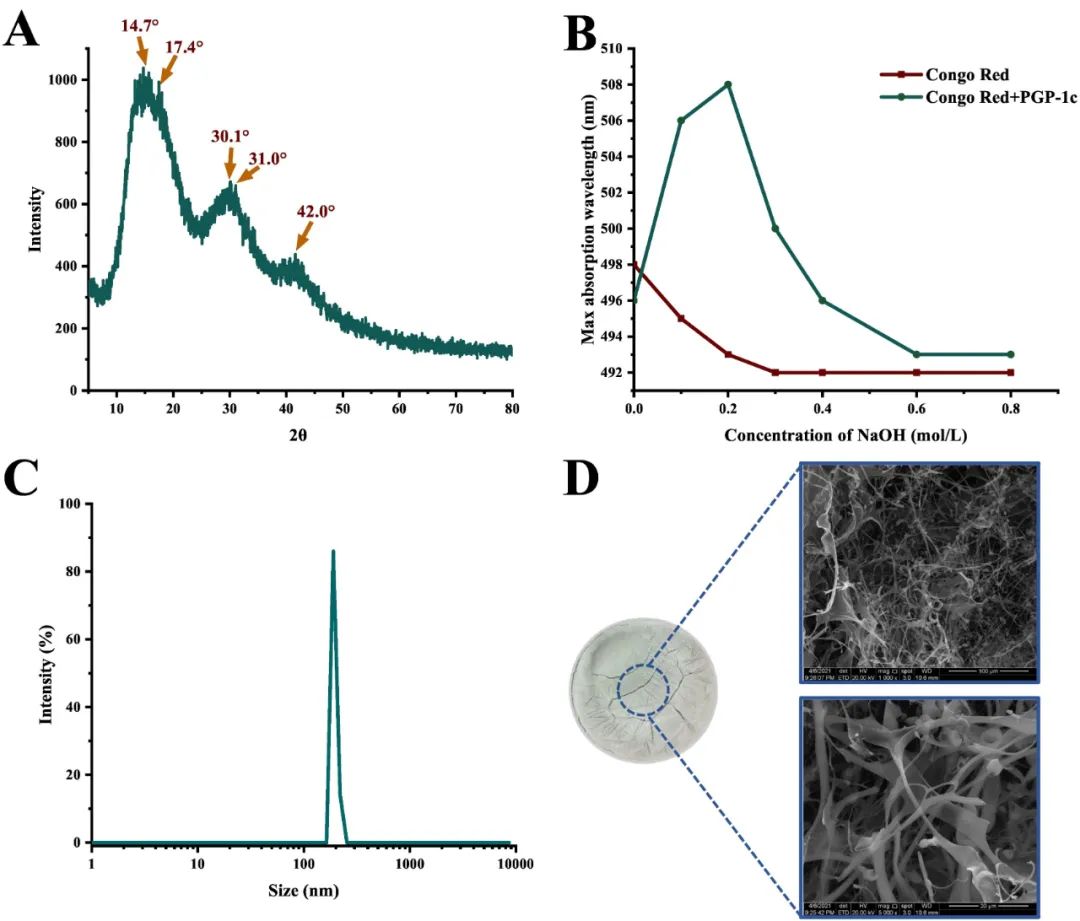
图4 PGP-1c的XRD图谱(A),三螺旋构象分析(B);通过动态光散射(C)进行尺寸分布;100 μm和20 μm(D)比例尺中的SEM图像
PGP-1c对RAW264.7细胞生存能力的影响
为了探讨PGP-1c在处理24 h后对RAW264.7细胞活力的影响,采用MTT方法评估PGP-1c在12.5~800 μg/mL剂量范围内的影响。结果(图5A)显示,在12.5~400 μg/mL的范围内,PGP-1c对RAW264.7细胞没有细胞毒性作用。根据结果和与文献的比较,选择了100、200和400 μg/mL的浓度进行后续研究。
PGP-1c的免疫增强作用
被激活的巨噬细胞可以产生一系列的趋化因子和细胞因子,在激活机体的适应性免疫反应和调节各种免疫反应中起着重要作用。NO可以作为一种关键的细胞内信使分子释放出来,使巨噬细胞能够有效地检测癌症和其他目标细胞,达到治疗目的。IL-6是一种主要由巨噬细胞和T细胞产生的细胞因子,有助于从先天免疫到后天免疫的转变。TNF-α主要由活化的巨噬细胞产生,是主要的免疫调节因子。它可以启动免疫调节,促进免疫细胞的活性,具有许多重要的生物效应,如抗恶性肿瘤、血液凝固和造血功能。
在图5B中,PGP-1c在100~400 μg/mL的浓度下可以明显刺激RAW264.7细胞分泌NO,并呈现出明显的剂量—效应关系。用100 μg/mL PGP-1c刺激细胞24 h后,NO分泌水平达到(12.03±0.13)μmol/L,比空白对照组多3.08 μmol/L,是LPS组的54.45%,均达到显著性差异(P<0.05)。PGP-1c可能有很好的免疫刺激作用,以及抑制肿瘤细胞增殖的能力,这一点从NO分泌的大幅度增加可以看出。图5C中,与空白对照组相比,不同浓度(200~400 μg/mL)的PGP-1c可以明显促进RAW264.7细胞中IL-6的分泌,并呈现出明显的剂量效应。在图5D中,PGP-1c(100~400 μg/mL)能明显促进RAW264.7细胞释放TNF-α,100和200 μg/mL的PGP-1c极显著(P<0.001)。PGP-1c促进RAW264.7细胞释放细胞因子的结果表明,PGP-1c具有良好的免疫增强能力。
对杏鲍菇的3-O-甲基化杂半乳糖的研究发现,3-O-杂半乳糖的存在可能是多糖免疫活性的关键因素。此外,影响食用菌结构免疫调节活性关系的因素是多样而复杂的。其中,三螺旋结构和叠层多链多糖具有良好的免疫学活性。PGP-1c都具有上述结构,因此推断PGP-1c的免疫活性与3-O-CH3基团、三螺旋结构和支链堆积结构密切相关。但是,PGP-1c的具体结构—活性关系和免疫调节机制还需要进一步研究。
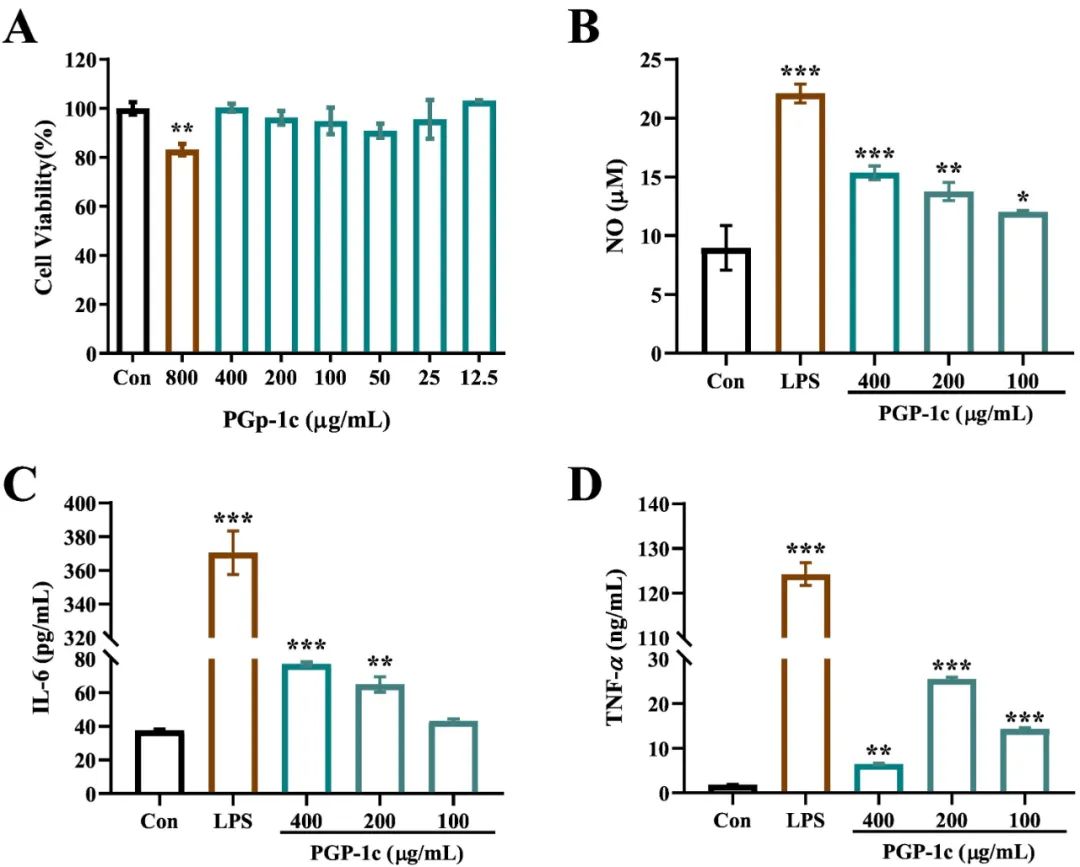
图5 PGP-1c对RAW264.7细胞活力的影响(A)。PGP-1c对RAW264.7细胞NO(B)、IL-6(C)和TNF-α(D)产生的影响
Conclusion
在这项工作中,从秀珍菇中获得的PGP-1c(20.9 kDa)的结构阐明表现为一种葡甘露半乳聚糖,由未取代的(1,6-α-Gal和1,6-α-3-OMe-Gal)和单取代的(1,2,6-α-Gal和1,2,6-α-3-OMe-Gal)半乳糖单位组成,通过α-(1→6)糖苷键作为骨架,可能的分支是由→3)-Glc-(1→Man和一些岩藻糖组成的长支链在半乳糖的O-2处连接,分支度为48%。此外,通过分子链的构象行为,观察到PGP-1c有一个堆积的大分子网络和三螺旋结构。此外,PGP-1c能以剂量依赖的方式大大增加NO和细胞因子的释放,表明其具有积极的免疫增强作用。然而,还需要对溶液特性和结构—活性联系进行更多的研究。PGP-1c良好的免疫学活性被认为与O-CH3基团和长支链有关,这为下一步的结构—功能关系和机制研究奠定了基础。